



Fungi
Eumycota: mushrooms, sac fungi, yeast, molds, rusts, smuts, etc.
Meredith Blackwell, Rytas Vilgalys, Timothy Y. James, and John W. Taylor
Click on an image to view larger version & data in a new window
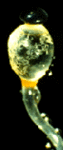




 This tree diagram shows the relationships between several groups of organisms.
This tree diagram shows the relationships between several groups of organisms.The root of the current tree connects the organisms featured in this tree to their containing group and the rest of the Tree of Life. The basal branching point in the tree represents the ancestor of the other groups in the tree. This ancestor diversified over time into several descendent subgroups, which are represented as internal nodes and terminal taxa to the right.
 You can click on the root to travel down the Tree of Life all the way to the root of all Life, and you can click on the names of descendent subgroups to travel up the Tree of Life all the way to individual species.
You can click on the root to travel down the Tree of Life all the way to the root of all Life, and you can click on the names of descendent subgroups to travel up the Tree of Life all the way to individual species.For more information on ToL tree formatting, please see Interpreting the Tree or Classification. To learn more about phylogenetic trees, please visit our Phylogenetic Biology pages.
close box
Phylogeny modified from James et al., 2006a, 2006b; Liu et al., 2006; Seif et al., 2005; Steenkamp et al., 2006.
Containing group: Eukaryotes
Introduction
The organisms of the fungal lineage include mushrooms, rusts, smuts, puffballs, truffles, morels, molds, and yeasts, as well as many less well-known organisms (Alexopoulos et al., 1996). More than 70,000 species of fungi have been described; however, some estimates of total numbers suggest that 1.5 million species may exist (Hawksworth, 1991; Hawksworth et al., 1995).As the sister group of animals and part of the eukaryotic crown group that radiated about a billion years ago, the fungi constitute an independent group equal in rank to that of plants and animals. They share with animals the ability to export hydrolytic enzymes that break down biopolymers, which can be absorbed for nutrition. Rather than requiring a stomach to accomplish digestion, fungi live in their own food supply and simply grow into new food as the local environment becomes nutrient depleted.
Most biologists have seen dense filamentous fungal colonies growing on rich nutrient agar plates, but in nature the filaments can be much longer and the colonies less dense. When one of the filaments contacts a food supply, the entire colony mobilizes and reallocates resources to exploit the new food. Should all food become depleted, sporulation is triggered. Although the fungal filaments and spores are microscopic, the colony can be very large with individuals of some species rivaling the mass of the largest animals or plants.
Click on an image to view larger version & data in a new window
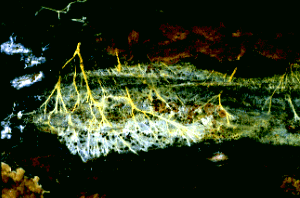
Figure 1: Hyphae of a wood-decaying fungus found growing on the underside of a fallen log. The metabolically active hyphae have secreted droplets on their surfaces. Copyright © M. Blackwell 1996.
Prior to mating in sexual reproduction, individual fungi communicate with other individuals chemically via pheromones. In every phylum at least one pheromone has been characterized, and they range from sesquiterpines and derivatives of the carotenoid pathway in chytridiomycetes and zygomycetes to oligopeptides in ascomycetes and basidiomycetes.Within their varied natural habitats fungi usually are the primary decomposer organisms present. Many species are free-living saprobes (users of carbon fixed by other organisms) in woody substrates, soils, leaf litter, dead animals, and animal exudates. The large cavities eaten out of living trees by wood-decaying fungi provide nest holes for a variety of animals, and extinction of the ivory billed woodpecker was due in large part to loss, through human activity, of nesting trees in bottom land hardwoods. In some low nitrogen environments several independent groups of fungi have adaptations such as nooses and sticky knobs with which to trap and degrade nematodes and other small animals. A number of references on fungal ecology are available (Carroll and Wicklow, 1992; Cooke and Whipps, 1993; Dix and Webster, 1995).
However, many other fungi are biotrophs, and in this role a number of successful groups form symbiotic associations with plants (including algae), animals (especially arthropods), and prokaryotes. Examples are lichens, mycorrhizae, and leaf and stem endophytes. Although lichens may seem infrequent in polluted cities, they can form the dominant vegetation in nordic environments, and there is a better than 80% chance that any plant you find is mycorrhizal. Leaf and stem endophytes are a more recent discovery, and some of these fungi can protect the plants they inhabit from herbivory and even influence flowering and other aspects of plant reproductive biology. Fungi are our most important plant pathogens, and include rusts, smuts, and many ascomycetes such as the agents of Dutch elm disease and chestnut blight. Among the other well known associations are fungal parasites of animals. Humans, for example, may succumb to diseases caused by Pneumocystis (a type of pneumonia that affects individuals with supressed immune systems), Coccidioides (valley fever), Ajellomyces (blastomycosis and histoplasmosis), and Cryptococcus (cryptococcosis) (Kwon-Chung and Bennett, 1992).
Click on an image to view larger version & data in a new window
Figure 2: The fluffy white hyphae of the mycorrhizal fungus Rhizopogon rubescens has enveloped the smaller roots of a Virginia pine seedling. Note that some of the mycelium extends out into the surrounding environment. Copyright © J. B. Anderson 1996.
Click on an image to view larger version & data in a new window

Figure 3: Entomophthora, "destroyer of insects", is the agent of a fungual infection that kills flies. After their death the fungal growth erupts through the fly cuticle, and dispersal by forcible spore discharge is a source of inoculum for infection of new flies. Copyright © G. L. Barron 1996.
Fungal spores may be actively or passively released for dispersal by several effective methods. The air we breathe is filled with spores of species that are air dispersed. These usually are species that produce large numbers of spores, and examples include many species pathogenic on agricultural crops and trees. Other species are adapted for dispersal within or on the surfaces of animals (particularly arthropods). Some fungi are rain splash or flowing water dispersed. In a few cases the forcible release of spores is sufficient to serve as the dispersal method as well. The function of some spores is not primarily for dispersal, but to allow the organisms to survive as resistant cells during periods when the conditions of the environment are not conducive to growth.Fungi are vital for their ecosystem functions, some of which we have reviewed in the previous paragraphs. In addition a number of fungi are used in the processing and flavoring of foods (baker's and brewer's yeasts, Penicillia in cheese-making) and in production of antibiotics and organic acids. Other fungi produce secondary metabolites such as aflatoxins that may be potent toxins and carcinogens in food of birds, fish, humans, and other mammals.
A few species are studied as model organisms that can be used to gain knowledge of basic processes such as genetics, physiology, biochemistry, and molecular biology with results that are applicable to many organisms (Taylor et al., 1993). Some of the fungi that have been intensively studied in this way include Saccharomyces cereviseae, Neurospora crassa, and Ustilago maydis.
Most phyla appear to be terrestrial in origin, although all major groups have invaded marine and freshwater habitats. An exception to this generality is the flagellum-bearing phyla Chytridiomycota, Blastocladiomycota, and Neocallimastigomycota (collectively referred to as chytrids), which probably had an aquatic origin. Extant chytrid species also occur in terrestrial environments as plant pathogenic fungi, soil fungi, and even as anaerobic inhabitants of the guts of herbivores such as cows (all Neocallimastigomycota).
Characteristics
Fungi are characterized by non-motile bodies (thalli) constructed of apically elongating walled filaments (hyphae), a life cycle with sexual and asexual reproduction, usually from a common thallus, haploid thalli resulting from zygotic meiosis, and heterotrophic nutrition. Spindle pole bodies, not centrioles, usually are associated with the nuclear envelope during cell division. The characteristic wall components are chitin (beta-1,4-linked homopolymers of N-acetylglucosamine in microcrystalline state) and glucans primarily alpha-glucans (alpha-1,3- and alpha-1,6- linkages) (Griffin, 1994).Click on an image to view larger version & data in a new window
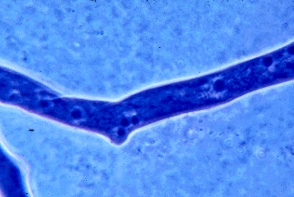
Figure 4: Portion of a hypha of a zygomycete stained with a blue dye to show the many nuclei present. Many other fungi have septations that devide the hyphae into compartments that usually contain one to several nuclei per compartment. Copyright © M. Blackwell 1996.
Click on an image to view larger version & data in a new window
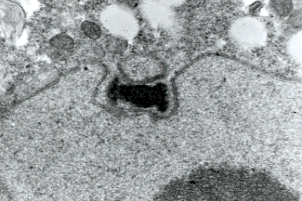
Figure 5: Transmission electron micrograph showing duplicated spindle pole body of a prophase I meiotic nucleus of a basidiomycete Exobasidium. Only chytrids among fungi have centrioles and lack spindle pole bodies. Copyright © Beth Richardson 1996.
Exceptions to this characterization of fungi are well known, and include the following: Most species of chytrids have cells with a single, smooth, posteriorly inserted flagellum at some stage in the life cycle, and centrioles are associated with nuclear division. The life cycles of most chytrids are poorly studied, but some (Blastocladiomycota) are known to have zygotic meiosis (therefore, alternation between haploid and diploid generations). Certain members of Mucoromycotina, Ascomycota, and Basidiomycota may lack hyphal growth during part or all of their life cycles, and, instead, produce budding yeast cells. Most fungal species with yeast growth forms contain only minute amounts of chitin in the walls of the yeast cells. A few species of Ascomycota (Ophiostomataceae) have cellulose in their walls, and certain members of Blastocladiomycota and Entomophthoromycotina lack walls during part of their life cycle (Alexopoulos et al., 1996).Fossil Record
Based on the available fossil record, fungi are presumed to have been present in Late Proterozoic (900-570 mya). Terrestrial forms of purported ascomycetes are reported in associations with microarthropods in the Silurian Period (438-408 mya) (Sherwood-Pike and Gray, 1985). Fossil hyphae in association with wood decay and fossil chytrids and Glomales-Endogonales representatives associated with plants of the Rhynie Chert are reported from the Devonian Period (408-360 mya) (Hass et al., 1994; Remy et al., 1994a, 1994b; Taylor et al., 1994a, 1995b). Fungal fossil diversity increased throughout the Paleozoic Era (Taylor et al., 1994b) with all modern classes reported in the Pennsylvanian Epoch (320-286 mya).A first attempt to match molecular data on fungal phylogeny to the geological record shows general agreement, but does point out some conflicts between the two types of data (Berbee and Taylor 1993).
Biogeography
Wherever adequate moisture, temperature, and organic substrates are available, fungi are present. Although we normally think of fungi as growing in warm, moist forests, many species occur in habitats that are cold, periodically arid, or otherwise seemingly inhospitable. It is important to recognize that optimum conditions for growth and reproduction vary widely with fungal species. Diversity of most groups of fungi tends to increase in tropical regions, but detailed studies are only in their infancy (Isaac et al., 1993).Although many saprobic and plant pathogenic species with low substrate specificity and effective dispersal systems have broad distributions, gene flow appears to be restricted in many fungi. For these species large bodies of water such as the Atlantic and Pacific Oceans create barriers to gene exchange. Some distributions are limited by substrate availability, and dramatic examples come from parasites of Gondowanan plants; one of these is the Southern Hemisphere distribution of the ascomycete Cyttaria, corresponding with part of the distribution of its host plant Nothofagus. The fossil record shows that fungi were present in Antarctica, as is the case for other organisms with Gondwanan distributions. Arthropod associates also may show distributions throughout part or all of a host range, and some fungal species (ex. wood wasp associates) occur outside the range of the associated arthropod.
Notable Fungi
- The largest known basidioma (mushroom or fruiting body) was that of a Rigidioporus ulmarius (Agaricomycetes), hidden-away in a shady corner of the Royal Botanic Gardens, Kew, Richmond, Surrey, England. This fruiting body was mentioned in the Guinness Book of World Records (Matthews, 1994). At the beginning of every New Year the Annual Mensuration Ceremony of the fruiting body took place, and on 19 January 1996 it had increased to 170 cm maximum length (up from 159 in 1995) and 146 cm maximum width (up from 140 in 1995). It also grew 4 cm taller from the soil level, measuring 54 cm. The weight of the fruiting body was estimated to be 284 kg (625 pounds)! Amid rumors of its destruction, Dr. Brian M. Spooner, Head of Mycology, Royal Botanic Gardens, has brought us up to date on the fate of the record specimen. Unfortunately, the basidioma began to rot at the edges a few years ago, likely because the hyphal body of the fungus digested away its elm root substrate, reminding us that a fungus needs a good dispersal system to escape the substrate that eventually inevitably is destroyed. In the life of the fruiting body many trillions of spores must have been produced, and some of these surely fell on an appropiate substrate to establish a new infection. The final insult to the fruiting body came from a fox that burrowed under one side and caused it to collapse.Other large basidiomata are those of a Canadian puffball almost 9 feet in circumference (over 48 pounds) and a basidiocarp of the sulfur mushroom in England (100 pounds). A previous Guinness Book of World Records record-sized fruiting body of Bridgeoporus nobilissimus, an endangered species of the Pacific Northwest of the United States, is over 160 kg (300 pounds) and may have regained the title of “largest” with the demise of the Kew specimen. This polypore also may do itself in because its great weight is likely to eventually cause it to fall as the mycelium depletes its food source, often the noble fir tree.Click on an image to view larger version & data in a new window
 Figure 6: The largest basidioma world record holder Rigidioporus ulmarius at Kew when it was still intact. The mushroom is shown in its largest dimension (170 cm or over 5 1/2 feet). Copyright © D. Pegler 1996.
Figure 6: The largest basidioma world record holder Rigidioporus ulmarius at Kew when it was still intact. The mushroom is shown in its largest dimension (170 cm or over 5 1/2 feet). Copyright © D. Pegler 1996. - Reproductive structures clearly can be very large, but what about the body of the fungus, which often is hidden from view within the substrate? One fungus body constructed of tubular filaments (hyphae) was brought to our attention when molecular techniques were used to show that it was extensive (37 acres and an estimated blue whale equivalent size of 110 tons). The Michigan fungus clone (Armillaria bulbosa, Agaricomycetes) grew in tree roots and soil. This report drew attention to an even larger fungal clone of Armillaria ostoyae, reported earlier in the state of Washington, which covered over 1,500 acres. Each clone began from the germination of a single spore over a thousand years ago. Although they probably have fragmented and are no longer continuous bodies, such organisms give us cause to think about what constitutes an individual.
- Penicillium chrysogenum (Ascomycota) is known for its production of the antibiotic penicillin. Although other antibiotics are produced by a variety of organisms, penicillin was the first to be developed. In the spring of 1996 a long dried out culture of the original isolate prepared by its discoverer, Sir Alexander Fleming in the late 1920s, was auctioned by Sotheby's of London and sold to a pharmaceutical company for 23,000 pounds. This price is insignificant when one considers the worth of this fungus, not only in sales of penicillin, but in terms of illnesses cured and lives saved. In the past a simple scratch or blister sometimes could result in a fatal infection such as the blister that resulted in the death of John Calvin Coolidge (1918-1924), the son of a U. S. president. However, misuse of penicillin and other antibiotics has resulted in selection of resistant microorganisms, and the threat of untreatable bacterial infections and diseases (for example Staphylococcus aureus and Escherichia coli and tuberculosis and syphilis) is still present in our homes and recreation areas.
- Fungal spores fill the air we breathe. On many days in some localities the number of fungal spores in the air far exceeds the pollen grains. Fungal spores also cause allergies; however, unlike seasonal pollen production, some fungi can produce spores all year long. The largest number of fungal spores ever sampled was over 5.5 million per cubic foot in Wales (Matthews, 1994).
- Basidiomycetes have always attracted a lot of attention because some of them have large basidiocarps, but the realization that all fungi are important in ecosystem function has drawn more attention to microscopic forms as well. For example a report on the secret sex life of a yeast-like ascomycete human pathogen, Coccidioides immitis, made a headline of the New York Times (6 February 1996, p. B7). This fungus causes Valley Fever and is endemic in parts of the southwestern United States. Although no one has been able to observe sexual reproduction in this species, molecular studies show genetic diversity that is best explained by occurrence of sexual reproduction in the life cycle.
- Another yeast-like ascomycete reported in the Dallas Morning News (28 August 1995, p. 8D) lives in the gut of cigar beetles and is essential to the beetle's health. Without the gut fungi to detoxify the plant material of toxins, the beetles would be poisoned. Keep on the lookout for other reports of fascinating fungal feats.
Discussion of Phylogenetic Relationships
The kingdom Fungi is a diverse clade of heterotrophic organisms that shares some characters with animals such as chitinous structures, storage of glycogen, and mitochondrial codon UGA encoding tryptophan. Both animals and fungi have spores or gametes with a single smooth, posteriorly inserted flagellum, but only species of the basal chytrid phyla have retained this primitive character (Barr, 1992; Cavalier-Smith, 1987, 1995). Fungi, animals, and other heterotrophic protist-like organisms such as choanoflagellates and Mesomycetozoea are now considered part of the larger group termed opisthokonts (Cavalier-Smith, 1987) in reference to the posterior flagellum.The branch uniting the fungi and animals is well-supported based on a number of molecular phylogenetic datasets, including the nuclear small subunit ribosomal RNA gene (Wainwright et al., 1993; Bruns et al. 1993), unique and shared sequence insertions in proteins such as elongation factor 1α (Baldauf and Palmer, 1993), entire mitochondrial genomes (Lang et al., 2002), and concatenated protein-coding genes (Steenkamp et al., 2006).
Prior classification systems of Fungi based primarily on morphology are in need of updating to more accurately reflect phylogenetic relationships as determined by molecular systematics. Molecular characters have been essential for phylogenetic analysis in cases when morphological characters are convergent, reduced, or missing among the taxa considered. This is especially true of species that never reproduce sexually, because characters of sexual reproduction traditionally have been the basis for classification of Fungi. Use of molecular characters allows asexual fungi to be placed among their closest relatives.
Previous classifications placed early-diverging fungal groups (non-Ascomycota or Basidiomycota) into two phyla: Chytridiomycota and Zygomycota. Numerous phylogenetic studies now suggest that neither is monophyletic, and the latest classification scheme includes six phyla and an additional four unplaced subphyla (Hibbett et al., 2007). At present, because of the ancient divergence times between the fungal phyla, the exact phylogenetic relationships are ambiguous. Chytrids appear to be a paraphyletic group at the base of the fungal phylogeny and merely fungal lineages which have retained the character of flagellated spores. Three phyla of flagellated fungi are proposed (Blastocladiomycota, Chytridiomycota, and Neocallimastigomycota; Hibbett et al., 2007) and two chytrid genera Olpidium and Rozella, are of uncertain phylogenetic position (James et al., 2006a, 2006b). These genera are interesting because they are both highly reduced endoparasites (living inside the host cell) whose entire thallus consists of only a spherical body absorbing nutrients from the host material that surrounds it. Rozella appears in an isolated position in the fungal phylogeny as the very earliest lineage to diverge from the rest of the fungi (James et al., 2006a, 2006b). In contrast, Olpidium brassicae appears to have diverged after the majority of chytrids and is more closely related to some zygomycete fungi (James et al., 2006a, 2006b).
Click on an image to view larger version & data in a new window
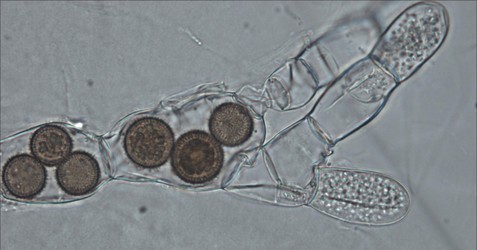
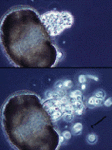
Figure 7: The endoparasitic chytrid Rozella allomycis inside the hyphae of another chytrid Allomyces. Thick spiny spores of the parasite are seen inside some cells while zoospores are produced in other cells. © Timothy Y. James
Evidence from shared morphological characters such as regularly septate hyphae and a dikaryotic stage (two separate and different nuclei in a single hyphal segment) in the life cycle usually has been interpreted as support for a close relationship between Basidiomycota and Ascomycota. Numerous phylogenetic studies such as SSU rDNA (Berbee and Taylor, 1992), RNA polymerase genes (Liu et al., 2006), and mitochondrial genome sequencing (Seif et al., 2005) provide strong support for this relationship. A subkingdom termed Dikarya is proposed (Hibbett et al., 2007), creating a division between a highly speciose subkingdom (Dikarya) and the remaining early diverging lineages whose relationships are not precisely known.
Fungal classification is far from static, and even which organisms are actually members of Fungi is changing. For example, the group trichomycetes describes gut inhabitants of arthropods that share similarities with zygomycetes. Molecular phylogenetic studies have demonstrated that two of the four orders of trichomycetes are actually members of the Mesomycetozoea protist group (Benny and O’Donnell, 2000; Cafaro, 2005). Other organisms that were previously considered to be Fungi because of their heterotrophic, mold-like growth forms are now classified as stramenopiles (Oomycota, Hyphochytriomycota, and Labyrinthulomycota) or slime molds (Myxomycota, Plasmodiomycota, Dictyosteliomycota, Acrasiomycota) (Bhattacharya et al., 1992; Leipe et al., 1994; Van der Auwera et al., 1995). More interesting for mycologists are the findings that some species previously considered protozoa are actually Fungi. For example, the species Hyaloraphidium curvatum was assumed to be a green alga that had adopted a heterotrophic lifecycle concomitantly with losing its chloroplast. It is now known to be a chytrid fungus related to Monoblephariomycetes but lacking a flagellated stage (Ustinova et al., 2000). Other examples include the parasitic organisms presumed to be protozoa, such as the cockroach parasite Nepridiophaga (Wylezich et al., 2004) and the Daphnia parasite Polycarum (Johnson et al., 2006) recently demonstrated to be members of the fungal kingdom based on SSU rDNA phylogenies.
The most revolutionary addition to the fungal lineage has occurred with phylogenetic evidence indicating the protist group microsporidia is closely related to Fungi–possibly derived from zygomycetes (Keeling, 2003) or sister to the genus Rozella on the earliest branch in the fungal kingdom (James et al., 2006a). Microsporidia are highly specialized intracellular parasites (primarily of animals) that lack mitochondria but have chitin and trehalose in their spores (similar to Fungi). All molecular studies have shown that microsporidia evolve at an extremely accelerated rate of evolution, making their placement in the Tree of Life difficult. The relationship with fungi is supported by many single and multiple gene phylogenies (e.g., Liu et al., 2006), but an exact placement within the fungi has not received strong support (Keeling and Fast, 2002).
More recently the nuclearid amoebae have been demonstrated to be a sister group to the Fungi with strong support (Steenkamp et al., 2006). This finding is significant because Nuclearia lacks a cell wall and has phagotrophic nutrition in which the food source (such as a bacterium or algal cell) is engulfed wholly, unlike fungi and microsporidia which utilize absorptive nutrition. Further sampling of basal fungal lineages will be needed to determine whether a Nuclearia-like organism was the cenancestor (most recent common ancestor) of Fungi.
0 comments:
Post a Comment